

Os óleos tóxicos que a medicina vem naturalizando
Óleos insaturados, tipo ômega 6 e ômega 3 – implicados em vários problemas metabólicos e doenças crônico-degenerativas


[Imagem: foodmatterslive.com]
Os óleos insaturados podem apresentar virtudes de algum tipo, no entanto jamais poderiam ser classificados como alimentos pró-metabólicos. Sequer como alimentos no melhor sentido da palavra.
E isso por uma simples razão: há várias gerações que se acumulam pesquisas científicas evidenciando os males desses óleos quimicamente instáveis, portadores daquelas cadeias de carbonos insaturados [C=C]. E inibidores da respiração celular [A95].
Tais óleos são procedentes da indústria de óleos de sementes, extraídos e depois encaminhados para a fabricação de lubrificantes, plásticos, tintas, cosméticos e alimentos [alimentos industriais utilizam cada vez menos gordura saturada e predominantemente óleos insaturados] [A1]
Na alimentação, como nós sabemos, as tradicionais banha de porco, a manteiga e a gordura animal - todas elas gorduras saturadas, saudáveis - em geral foram tendo seu uso desencorajado, para darem lugar aos óleos insaturados, tipicamente industriais. Não foi um processo adequadamente conduzido: os congressos médicos e faculdades de medicina, embalados na desinformação e propaganda industrial, não abriram suficiente debate sobre os óleos insaturados e a saúde humana. Assim o mito foi sendo construído.
Aqui se estabeleceu a seguinte contradição: mais artigos científicos revelando o papel tóxico desses óleos extraídos de sementes não foram traduzidos em mais esclarecimento para o público; claro, se a corporação médica é a primeira a naturalizar tais óleos, embalada por sua vez pela Big Mídia, como imaginar que a população possa encontrar a informação corretaɁ
Não encontra facilmente. E nem à primeira vista. A mitologia opera como cortina de fumaça [de desinformação, diriam hoje].
E é por isso que o pensamento hegemônico sobre óleos como girassol, milho, soja, amendoim, amêndoas, linhaça [A2] e afins continua acolhendo tais produtos como “alimentícios”, quando, no real, são tóxicos daqueles que impactam negativamente os nossos vários sistemas orgânicos além de ficarem armazenados no nosso tecido adiposo por longuíssimos anos.
Existe um conhecimento comum ou senso comum sobre os diferentes tipos de gorduras que é o que termina prevalecendo na televisão, nos sites de “boa saúde” e até na opinião de doutores. “A influência das indústrias médica e farmacêutica é tão pervasiva que [aquele senso comum sobre as gorduras, GMF] termina se tornando o pressuposto da maioria das pesquisas biológicas” [R. Peat].
Ao armar sua pesquisa, o cientista costuma partir do princípio – mitológico – de que gordura saturada faz mal, é patogênica. Dá doença cardiovascular.
“Felizmente muita gente está trabalhando por fora do mito, em problemas especializados da fisiologia e da biologia celular e suas observações estão mostrando a realidade como algo muito mais interessante e complexo do que a mitologia” [R. Peat].
O mito costuma deixar de levar em consideração que “os óleos insaturados, sintetizados pelas plantas [no caso dos óleos de peixe, eles são sintetizados por algas] são menos estáveis que as gorduras saturadas, e os ômega 3 e ômega 6, derivados das plantas são muito susceptíveis a serem quebrados em toxinas, especialmente em animais de sangue quente” [R. Peat].

[Imagem: actinutrition.fr\sante]
Uma diferença entre os insaturados e os saturados é que aqueles apresentam propriedades patogênicas também pelo fato de serem um pouco mais solúveis em água que os saturados. “Os óleos insaturados são muitas vezes mais solúveis na água do que os gorduras saturadas do mesmo comprimento. Esta propriedade provavelmente explica porque apenas o ácido palmítico [saturado] funciona como um surfactante nos pulmões, permitindo que os alvéolos fiquem abertos, ao passo que os óleos insaturados causam edema pulmonar e falência respiratória” [R. Peat]. As óleos insaturados ativam mastócitos [A15]. Promovem crise respiratória aguda [A16].
Mas seu poder tóxico vai muito além.
Vejamos algumas informações do mundo científico – naturalmente não da ciência que é mais seletivamente divulgada – sobre tais óleos.
Em primeiro lugar sua instabilidade química: por sua estrutura molecular pobre em termos de saturação [seus carbonos não estão todos saturados com hidrogênio como na gordura saturada], são facilmente oxidáveis diante do oxigênio, exposição à luz, calor, inclusive já quando estão dentro da suas garrafas.
Ao serem consumidos promovem inflamação, inibição do metabolismo celular e envelhecimento. Então, sim, são “alimentos” pró-aging.
São imunossupressores. Óleo insaturado [DHA] é comprovadamente imunossupressor [A58], assim como os demais insaturados. As endotoxinas são tóxicas, mas sua toxicidade é potencializada pelos óleos insaturados [A5][A8].
Óleo insaturado inibe respiração mitocondrial [A17] [A19] E também promove peroxidação lipídica [A20] Óleo insaturado compromete o metabolismo hepático [A59].
Mesmo em pequena quantidade diária, consumo de óleo insaturado [EPA] claramente aumenta a coagulabilidade e atividade fibrinolítica do sangue [A70] Óleo insaturado ativa plaquetas [A81]. E intoxica o cérebro [A83]. Óleo insaturado – e seus metabólitos - promove edema cerebral [A29].
Óleo insaturado desencadeia processos inflamatórios [A34]. E está envolvido na gênese da enxaqueca [A35].
A serotonina libera óleos insaturados nos tecidos [A37]. O óleo insaturado ativa plaquetas [A38] e aumenta síntese de fator de necrose tumoral e interleucina-6 [A39]; portanto age como inflamatório. Tumoração, aterosclerose e hiperinsulinemia foram demonstrados em pessoas consumidoras do ômega 6 [A22]
Membranas celulares de ratos com menor longevidade são peroxidadas com óleos insaturados [A63]. Ômega 3 é tóxico para o coração [A69]. Óleo insaturado faz parte da construção da placa ateromatosa arterial [A41]. E intoxica o músculo cardíaco [A46]. Óleos insaturados constróem aterosclerose [A74].

[Imagem: supaexport.ro]
Oferta de ômega 3 [principalmente DHA] aumenta susceptibilidade das membranas celulares à peroxidação lipídica e promove disrupção do sistema antioxidante [A72].
Ômega 3 aumenta insulinemia e promove acumulação severa de gordura marrom no fígado dos animais [A73]. Ômega 3 envelhece hemácias [A91] e promove câncer de mama [A99]. Os óleos insaturados tipo ômega 3 possuem impacto direto no desenvolvimento da resistência insulínica e diabetes [A21]. Ômega 3 induz ativação plaquetária [aciona fator de ativação plaquetária, FAP]. Ômega 3 promove peroxidação lipídica [A48], assim como os ômegas 6.
Obesidade pode ser promovida pelos óleos insaturados [A18]. Obesidade e óleos insaturados, uma relação patogênica: “níveis aumentados de metabólitos de ácidos graxos insaturados podem estar envolvidos na aumentada produção de cortisol observada em humanos obesos” [A28]. Óleos insaturados além de impulsionarem a obesidade [A60], promovem a osteoporose [A40]. A construção das manchas da velhice [lipofucsina] está vinculada aos óleos insaturados peroxidados [A64][A82]. Acúmulo de óleos insaturados na retina com o envelhecimento vem lado a lado com mais presença de radicais livres e seus danos [A25]
Em animais já ficou bem claro: o grau de instauração dos óleos nos tecidos está inversamente correlacionado com sua longevidade, isto é, mais óleo insaturado, menos longevidade consegue o animal [A3][A12].
Homens com Alzheimer possuem níveis mais altos de estrogênio que homens normais. Pois bem, a quantidade de DHA [ácido docosa-hexaenoico, do tipo ômega 3] no cérebro e em outros tecidos aumenta com o envelhecimento e seus produtos de fragmentação, incluindo neuroprostanos, estão associados com a demência. Níveis mais altos de DHA e de óleo insaturado total são encontrados no plasma de pacientes com demência, como explica R. Peat.
As gorduras saturadas são, ao contrário dos óleos insaturados, terapêuticas [A11][A9]. “Os gorduras saturadas possuem um efeito supressivo no aumento do CRH-induzido pelo ACTH, em pituitária de ratos”, ou seja, gordura saturada tem efeito antiinflamatório [A27][A96][A100].
Os efeitos protetores para a saúde de parte das gorduras saturadas foi mais de uma vez demonstrado. “Uma das mais claras evidências dos efeitos protetores das gorduras saturadas foi publicado pelo grupo de A A Nanji [A89] mostrando que a gordura saturada pode reverter inflamação, necrose e fibrose da doença alcoólica do fígado, mesmo quando a pessoa continua consumindo álcool, ao passo que o óleo de peixe e outros insaturados exacerbam o problema” [R. Peat].

[Imagem: mindovermunch.com]
Quando a pessoa entra em estresse, a presença de óleo insaturado nos tecidos do corpo torna o estresse muito mais perigoso e patogênico. “Estresse [que aumenta o pool de ácidos graxos livres e baixa a energia celular] torna a célula mais permeável, admitindo a entrada de amplo leque de substâncias, incluindo aquelas menos lipofílicas” [R. Peat] como é o caso dos óleos insaturados.
Estes se ligam a estruturas celulares, aumentando sua afinidade pela água, promovendo edema generalizado, inclusive nas mitocôndrias, fazendo com que mais cálcio entre na célula, ativando processos excitatórios, girando o sistema de oxidação para inflamação, com possível crescimento inapropriado da célula e até sua morte.
Apesar de sua instabilidade, de sua tendência a se quebrar em fragmentos tóxicos e serem os ácidos graxos preferencialmente liberados dos adipócitos durante o estresse, a proporção de óleos insaturados nos tecidos aumenta com a idade.
Um motivo é que as gorduras saturadas são preferencialmente oxidadas por muitos tipos de células. A albumina preferencialmente entrega nas células a gordura saturada em vez do óleo insaturado.
“Ao mesmo tempo em que a competição do óleo insaturado por sites de ligação de proteínas bloqueia o efeito do hormônio tireoidiano e da vitamina A, a ação do óleo insaturado nas proteínas carreadoras de hormônios esteroidais sexuais como SBP ou SSBG [sex steroid binding globulin, isto é, a globulina que faz esse papel carreador] aumenta a atividade do estrogênio” [R. Peat].
Ácidos graxos insaturados livres impedem que o estrogênio se ligue àquela proteína, aumentando a atividade estrogenizante dessa forma. E é aquela proteína que mantém o estrogênio fora das células.
Portanto a relação entre os óleos insaturados e o estrogênio amplia o efeito nefasto da ação dos dois. Em português fluente: óleos insaturados são, em si mesmos, um poderoso agente estrogenizante.
O bebê nasce com quase nenhum óleo insaturado nos seus tecidos [A23] e com mitocôndrias ricas em gorduras saturadas em suas membranas; com o tempo, com a alimentação do bebê trazendo óleos insaturados - as malfadadas “papinhas” com soja - , a membrana mitocondrial vai degradando, vai substituindo a gordura saturada pela insaturada e deprimindo, ao longo dos anos, o seu metabolismo.
A presença de óleo insaturado nos tecidos adiposos das pessoas aumentou [A26] em mais de cem por cento no último meio século, o que está correlacionado com um aumento do consumo de tais óleos no mesmo período.
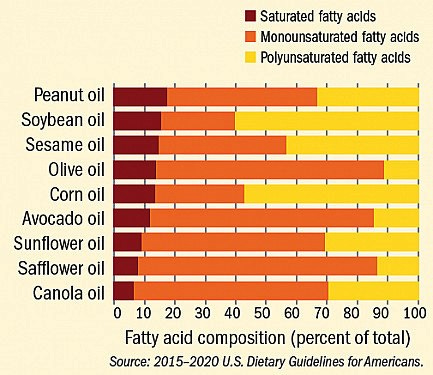
[Imagem: healthharvard.edu] Observe-se que o óleo de soja e o de milho disputam o primeiro lugar em ômegas.
À guisa de conclusão, pode-se dizer que o óleo insaturado é tóxico de várias formas e naquilo que ele porventura pudesse ser benéfico, tal efeito ficaria amplamente superado pela sua natureza inevitavelmente antimetabólica.
Óleo insaturado derruba o açúcar sanguíneo (danifica a mitocôndria, fazendo com que o combustível seja queimado sem o seu efeito pleno; suprime a atividade da enzima respiratória e a gira os açúcares para a produção de gordura; aliás, Por essa razão os óleos insaturados são usados em regime de engorda por criadores de porcos.
E são diabetogênicos.
Em suma, o óleo insaturado inibe o metabolismo ao mesmo tempo em que o açúcar sobrante vai ser convertido em gordura. O organismo que armazena óleos insaturados estará mais estressado (baixo metabolismo), mais tendente à obesidade.
Os óleos insaturados manterão o organismo sob o regime dos hormônios do estresse, do cortisol, também por conta das sucessivas quedas do açúcar plasmático que o óleo insaturado induz.
Em suma, fica difícil entender porque algum profissional de saúde recomendaria – conscientemente - um óleo com esse perfil de inibidor e agressor metabólico, claramente construtor do hipotireoidismo.
GM Fontes, Brasília, 12-4-23
As informações aqui presentes não pretendem servir para uso diagnóstico, prescrição médica, tratamento, prevenção ou mitigação de qualquer doença humana. Não pretendem substituir a consulta ao profissional médico ou servir como recomendação para qualquer plano de tratamento. Trata-se de informações com fins estritamente educativos.
Referências ___________________
[A1] Opinião da indústria - FATS AND OILS: Many Challenges Faced by Industry, Consumers. “The food industry has aggressively decreased the level of saturated fats in processed foods, but such a switch has not been easy since solid fats play an important role in the shelf-life and acceptability of foods.[...] For example, before the wide-spread availability of vegetable oils, most foods were produced with animal fats such as butter, lard and tallow (the original fat in fast food French fries). However, in the late 1950s and early 1960s, nutritionists thought that cholesterol consumption was related to heart disease so the food industry decreased the utilization of animal fats in many foods. Since solid fats play so many important roles in foods, something had to replace the animal fats and in this case it was mainly tropical oils (palm, coconut and palm kernel). Tropical fats are solid because they are also high in saturated fatty acids (e.g. coconut oil is 85% saturated fatty acids)”. References: 1. Vieira, S.A.; McClements, D.J.; Decker E.A. Challenges of utilizing healthy fats in foods. Adv Nutr. 2015 May 15;6(3):309S-17S. 2. Kritchevsky D. History of recommendations to the public about dietary fat. J Nutr. 1998; 128(2):449S-52S. Disponível em: https://www.soyconnection.com/resources/health-nutrition-newsletter/newsletter-article-list/fats-and-oils-many-challenges-faced-by-industry-consumers
[A2] Sobre o óleo de linhaça eis o que diz a propaganda da indústria de madeira: “O óleo de linhaça é usado há séculos para proteger e acabar madeira. É sem dúvida um dos acabamentos para madeira mais populares do mundo. Protege a madeira, seca lentamente e dá um acabamento quente com lustro discreto que acentua os veios da madeira”. https://ironfittings.com.br/produto/oleo-de-linhaca-refinado/
[A3] BARJA G, 2004. Trends Neurosci. 2004 Oct;27(10):595-600. Free radicals and aging. Barja G. "The degree of unsaturation of tissue fatty acids also correlates inversely with maximum longevity."
[A4] BORBA J M, 2010. Nutr Neurosci. 2010 Jun;13(3):144-50. Essential fatty acid deficiency reduces cortical spreading depression propagation in rats: a two-generation study. Borba JM, et al.
[A4] BRONNIKOV G E, 2010. Biochemistry (Mosc). 2010 Dec;75(12):1491-7. Dietary supplementation of old rats with hydrogenated peanut oil restores activities of mitochondrial respiratory complexes in skeletal muscles. Bronnikov GE, Kulagina TP, Aripovsky AV.
[A5] COOK, J A, 1979. Circ Shock. 1979;6(4):333-42. Resistance of essential fatty acid-deficient rats to endotoxic shock. Cook JA, Wise WC, Callihan CS.
[A6] CORONEL, R, WILM-SCHOPMAN F J, 2007. Cardiovasc Res. 2007 Jan 15;73(2):386-94. Dietary n-3 fatty acids promote arrhythmias during acute regional myocardial ischemia in isolated pig hearts. Coronel R, Wilms-Schopman FJ, Den Ruijter HM, Belterman CN, Schumacher CA, Opthof T, Hovenier R, Lemmens AG, Terpstra AH, Katan MB, Zock P.
[A7] GILTAY E J, GOOREN L J, 2004. Am J Clin Nutr. 2004 Nov;80(5):1167-74. Docosahexaenoic acid concentrations are higher in women than in men because of estrogenic effects. Giltay EJ, Gooren LJ, Toorians AW, Katan MB, Zock PL.
[A8] LI E J COOK J A, 1990. Circ Shock. 1990 Jun;31(2):159-70. Resistance of essential fatty acid-deficient rats to endotoxin-induced increases in vascular permeability. Li EJ, Cook JA, Spicer KM, Wise WC, Rokach J, Halushka PV.
[A9] NANJI A A JOKELAINAN K, 2001. J Pharmacol Exp Ther. 2001 Nov;299(2):638-44. Dietary saturated fatty acids reverse inflammatory and fibrotic changes in rat liver despite continued ethanol administration. Nanji AA, Jokelainen K, Tipoe GL, Rahemtulla A, Dannenberg AJ.
[A10] SERGEEVA M SISTEMA|ROKIN, M. 2005. Reprod Nutr Dev. 2005 Sep-Oct;45(5):633-46. Regulation of intracellular calcium levels by polyunsaturated fatty acids, arachidonic acid and docosahexaenoic acid, in astrocytes: possible involvement of phospholipase A2. Sergeeva M, Strokin M, Reiser G.
[A11] THOMSON A B, 1989. Am J Physiol. 1989 Jan;256(1 Pt 1):G178-87. Saturated fatty acid diet prevents radiation-associated decline in intestinal uptake. Thomson AB, Keelan M, Lam T, Cheeseman CI, Walker K, Clandinin MT.
[A12] VALENCAK T G, 2007. Aging Cell. 2007 Feb;6(1):15-25. N-3 polyunsaturated fatty acids impair lifespan but have no role for metabolism. Valencak TG, Ruf T.
[A13] Neurotoxicology. 2007 Nov;28(6):1220-9. Detrimental effects of post-treatment with fatty acids on brain injury in ischemic rats. Yang DY, Pan HC, Yen YJ, Wang CC, Chuang YH, Chen SY, Lin SY, Liao SL, Raung SL, Wu CW, Chou MC, Chiang AN, Chen CJ.
[A14] CHAN P H, FISHMAN R A, 1983. Induction of brain edema following intracerebral injection of arachidonic acid. Chan PH, Fishman RA, Caronna J, Schmidley JW, Prioleau G, Lee J.Ann Neurol. 1983 Jun;13(6):625-32. doi: 10.1002/ana.410130608.PMID: 6309072.
[A15] MASINI, E, PALMERANI, B, 1990.Biochem Pharmacol. 1990 Mar 1;39(5):879-89. Histamine release from rat mast cells induced by metabolic activation of polyunsaturated fatty acids into free radicals. Masini E, Palmerani B, Gambassi F, Pistelli A, Giannella E, Occupati B, Ciuffi M, Sacchi TB, Mannaioni PF.
[A16] BURSTEN S L, FEDERIGHI D AUTOR, 1996. Crit Care Med. 1996 Jul;24(7):1129-36. An increase in serum C18 unsaturated free fatty acids as a predictor of the development of acute respiratory distress syndrome. Bursten SL, Federighi DA, Parsons P, Harris WE, Abraham E, Moore EE Jr, Moore FA, Bianco JA, Singer JW, Repine JE.
[A17] COCCO, T, Di PAOLA M, 1999. Free Radic Biol Med. 1999 Jul;27(1-2):51-9. Arachidonic acid interaction with the mitochondrial electron transport chain promotes reactive oxygen species generation. Cocco T, Di Paola M, Papa S, Lorusso M. “It is shown that arachidonic acid causes an uncoupling effect under state 4 respiration of intact mitochondria as well as a marked inhibition of uncoupled respiration. While, under our conditions, the uncoupling effect is independent of the fatty acid species considered, the inhibition is stronger for unsaturated acids. Experiments carried out with mitochondrial particles indicated that the arachidonic acid dependent decrease of the respiratory activity is caused by a selective inhibition of Complex I and III. It is also shown that arachidonic acid causes a remarkable increase of hydrogen peroxide production when added to mitochondria respiring with either pyruvate+malate or succinate as substrate.” Citado por R. Peat.
[A18] GAIVA M H, COUTO R C, 2001. Br J Nutr. 2001 Sep;86(3):371-7. Polyunsaturated fatty acid-rich diets: effect on adipose tissue metabolism in rats. Gaiva MH, Couto RC, Oyama LM, Couto GE, Silveira VL, Riberio EB, Nascimento CM. “Wistar rats were fed ad libitum, for 8 weeks with one of the following diets: C, rat chow; S, rat chow containing 15 % (w/w) soyabean oil; F, rat chow containing 15 % (w/w) fish oil; SF, rat chow containing 15 % (w/w) soyabean and fish oil (5:1, w/w).” "Energy intake was reduced while carcass lipid content was increased in the three fat-fed groups." "These results indicate that enrichment of the diet with polyunsaturated fatty acids causes changes in adipose tissue metabolism that favour fat deposition. Different metabolic pathways were preferentially affected by each type of fatty acid used." Citado por R. Peat.
[A19] HERRERO A, 2001. Mech Ageing Dev 2001 Apr 15;122(4):427-43. Effect of the degree of fatty acid unsaturation of rat heart mitochondria on their rates of H2O2 production and lipid and protein oxidative damage. Herrero A, Portero-Otin M, Bellmunt MJ, Pamplona R, Barja G. "Previous comparative studies have shown that long-lived animals have lower fatty acid double bond content in their mitochondrial membranes than short-lived ones. In order to ascertain whether this trait protects mitochondria by decreasing lipid and protein oxidation and oxygen radical generation, the double bond content of rat heart mitochondrial membranes was manipulated by chronic feeding with semi-purified AIN-93G diets rich in highly unsaturated (UNSAT) or saturated (SAT) oils. UNSAT rat heart mitochondria had significantly higher double bond content and lipid peroxidation than SAT mitochondria. They also showed increased levels of the markers of protein oxidative damage malondialdehyde-lysine, protein carbonyls, and N(e)-(carboxymethyl)lysine adducts." "These results demonstrate that increasing the degree of fatty acid unsaturation of heart mitochondria increases oxidative damage to their lipids and proteins, and can also increase their rates of mitochondrial oxygen radical generation in situations in which the degree of reduction of Complex III is higher than normal. These observations strengthen the notion that the relatively low double bond content of the membranes of long-lived animals could have evolved to protect them from oxidative damage." Citado por R. Peat.
[A20] SONG J H, FUJIMOTO K, 2000. J Nutr 2000 Dec;130(12):3028-33. Polyunsaturated (n-3) fatty acids susceptible to peroxidation are increased in plasma and tissue lipids of rats fed docosahexaenoic acid-containing oils. Song JH, Fujimoto K, Miyazawa T.. "Thus, high incorporation of (n-3) fatty acids (mainly DHA) into plasma and tissue lipids due to DHA-containing oil ingestion may undesirably affect tissues by enhancing susceptibility of membranes to lipid peroxidation and by disrupting the antioxidant system." Citado por R. Peat.
[A21] STEERENBERG P A, BEEKHOF P K, 2002. Diabetes Nutr Metab. 2002 Aug;15(4):205-14. Long-term effect of fish oil diet on basal and stimulated plasma glucose and insulin levels in ob/ob mice. Steerenberg PA, Beekhof PK, Feskens EJ, Lips CJ, Hoppener JW, Beems RB. “We have investigated, in comparison to low and high fat diets, the effect of a fish oil diet on basal and stimulated plasma glucose and insulin levels in male and female ob/ob mice.” “Intercurrent deaths were found especially in the fish oil diet group. Compared to the other diet groups, plasma insulin levels of the fish oil diet group were significantly increased 3 months after the start of the diet and remained higher for another 3 months.” “At 12 months, microscopy revealed an increased severity of hepatic brown pigment accumulation and extramedullary haematopoiesis in the spleen of mice fed with fish oil.” “Fish oil diet also increased intercurrent mortality. However, a consistent course of death could not be established using morphological parameters.” Citado por R Peat.
[A22] YAM D, ELIRAZ A, 1996. Isr J Med Sci. 1996 Nov;32(11):1134-43. Diet and disease--the Israeli paradox: possible dangers of a high omega-6 polyunsaturated fatty acid diet. Yam D, Eliraz A, Berry EM. “Thus, rather than being beneficial, high omega-6 PUFA diets may have some long-term side effects, within the cluster of hyperinsulinemia, atherosclerosis and tumorigenesis.”
[A23] HORNSTRA G, 1990. Early Hum Dev. 1990 Dec;24(3):239-48.
Biochemical EFA status of mothers and their neonates after normal pregnancy.
Al MD, Hornstra G, van der Schouw YT, Bulstra-Ramakers MT, Huisjes HJ.
The essential fatty acid (EFA) status of neonates was compared with that of their mothers by determining the fatty acid compositions of phospholipids (PL), isolated from umbilical arterial and venous tissue, blood cells (BC) and plasma, from maternal venous plasma and BC, and from non-infarcted placental tissue. The PL of umbilical arterial tissue (efferent fetal vessels) contained fewer fatty acids of the (n-6) family and more of the (n-9) family than umbilical venous tissue (afferent fetal vessel). The relative amounts of (n-6) and (n-3) fatty acids were less in arterial than in venous plasma. Mead acid, 20:3(n-9), the presence of which indicates a poor EFA status, was 5 times higher in the efferent than in afferent cord vessels. In neonatal plasma and BC it was twice as high as compared with maternal levels. In general, the fatty acid composition of the placenta PL showed a comparable pattern as neonatal venous plasma PL. These findings demonstrate that the biochemical EFA status of neonates after a normal pregnancy is not optimal. The significant correlations between neonatal and maternal EFAs indicate that the neonatal EFA status depends on the EFA content of the maternal diet. Ou seja, o óleo insaturado da dieta da mãe dificilmente passa para os tecidos do bebê; este nasce com gorduras saturadas na sua membrana mitocondrial [cardiolipina, um fosfolipídeo que contém ácido palmítico]. À medida que a criança vai sendo alimentada com papinhas feitas com óleos insaturados, aquele ácido palmítico da cardiolipina vai sendo substituído por óleos insaturados, menos estáveis e menos capazes de suportar a atividade da enzima respiratória crucial, a citocromo oxidase, como explica R. Peat. Essa substituição da gordura saturada da mitocôndria por óleos insaturados, conforme a criança vai sendo alimentada erradamente, vai corresponder a um declínio da taxa metabólica daquela pessoa ao longo do tempo.
[A24] SMÍDOVÁ L, BASE J, 1990. Physiol Bohemoslov. 1990;39(2):125-34.
Proportion of individual fatty acids in the non-esterified (free) fatty acid (FFA) fraction in the serum of laboratory rats of different ages. Smídová L, Base J, Mourek J, Cechová I.
In experiments on Wistar strain rats of both sexes, aged 5, 10 and 14 days and adult (90-120 days), of their own breed, the authors determined the quantitative proportion of individual fatty acids in the serum free non-esterified fatty acid (FFA) fraction, using mixed blood (obtained by decapitation) and the titration method of Trout et al. (1960). The proportion of the individual fatty acids was then determined in this fraction by gas chromatography (Base 1978) and their concentration (in mumol.1-1) was determined by simple calculation from the relative chromatogram data. Animals in the first three age groups were killed in the morning, directly from the nest; in adult rats the FFA fraction was measured after a 20 h fast. It was demonstrated that the increase in the proportion of monoenoic acids was highly statistically significant (about fivefold) during ontogenesis and that there was also a marked increase in the quantitative expression of polyenic acids, especially in group (n-6). The n-6/n-3 acid index in the FFA fraction altered during maturation (despite some fluctuation it basically rose from 4.3 in 5-day-old young to 10.0 in adult rats). It was further demonstrated that the concentration of fatty acids with a very short chain fell significantly during development, so that C 8:0, for example, could be detected only in the first two age groups, but not in 14-day-old and adult rats. The concentration of the saturated fatty acids C 15:0 to C 18:0 in the serum FFA fraction showed a statistically significant increase, while the index expressing the ratio of saturated to unsaturated fatty acids displayed a downward trend during development. Citado por R. Peat.
[A25] NOUROOZ-ZADEH J, PEREIRA, P, 1999. Ophthalmic Res. 1999;31(4):273-9. Age-related accumulation of free polyunsaturated fatty acids in human retina.
Nourooz-Zadeh J, Pereira P. The present study reports composition of free (nonesterified) as well as total (sum of free and esterified) fatty acids (FAs) in human retina (n = 13). For free fatty acid (FFA) analysis, retina tissue was homogenized, total lipids were partitioned with ethyl acetate and subsequently applied onto a aminopropyl (NH2) cartridge to isolate FFAs from the bulk of other lipids. FFAs were converted to methyl ester derivatives and analysed by gas chromatography using flame ionization detector. Analysis of FFAs revealed that the mean percentage composition of the major components including palmitic acid (PA), stearic acid (SA), oleic acid (OA), arachidonic acid (AA) and docosahexaenoic acid (DHA) were 17.2, 36.7, 15.6, 8.8 and 14.2%, respectively. There were significant correlations between age of the donors’ and the content of both free AA and DHA (rPearson = 0.69, p = 0.005, and rPearson = 0.64, p = 0.009). The mean percentage of total PA, SA, OA, AA and DHA were 22.6, 23.2, 17.7, 11.4 and 21.9%, respectively. There was no association between age and any of the major FAs. The present study provides the first evidence for the presence of FFAs in the human retina as well as an age-related accumulation of polyunsaturated fatty acids (PUFAs). The latter finding suggests an alteration in the metabolism of retinal PUFAs which can be due to an increase of oxidative stress and/or decrease of antioxidant defences during ageing. Citado por R Peat.
[A26] STEPHAN J CARLSON, 2015. Adv Nutr November 2015 Adv Nutr vol. 6: 660-664, 2015 Increase in Adipose Tissue Linoleic Acid of US Adults in the Last Half Century. Stephan J Guyenet and Susan E Carlson. Linoleic acid (LA) is a bioactive fatty acid with diverse effects on human physiology and pathophysiology. LA is a major dietary fatty acid, and also one of the most abundant fatty acids in adipose tissue, where its concentration reflects dietary intake. Over the last half century in the United States, dietary LA intake has greatly increased as dietary fat sources have shifted toward polyunsaturated seed oils such as soybean oil. We have conducted a systematic literature review of studies reporting the concentration of LA in subcutaneous adipose tissue of US cohorts. Our results indicate that adipose tissue LA has increased by 136% over the last half century and that this increase is highly correlated with an increase in dietary LA intake over the same period of time.
[A27] KATOH K ASARI M, 2004. Comp Biochem Physiol A Mol Integr Physiol. 2004 Feb;137(2):357-64. Saturated fatty acids suppress adrenocorticotropic hormone (ACTH) release from rat anterior pituitary cells in vitro. Katoh K, Asari M, Ishiwata H, Sasaki Y, Obara Y. We studied whether fatty acids modify adrenocorticotropic hormone (ACTH) release induced by stimulation with corticotropin-releasing hormone (CRH) from rat anterior pituitary cells. Stimulation with CRH (0.01-100 nmol/l) significantly and concentration-dependently increased ACTH release, which was synergistically enhanced by the simultaneous stimulation with 1 nmol/l arginine-vasopressin. Addition of saturated fatty acids (butyrate, caprylate, laurate, palmitate and stearate) in a medium at 1 mmol/l, despite effects on the basal release, significantly reduced the ACTH release induced by CRH (1 nmol/l) stimulation. Caprylate suppressed ACTH release in a concentration-dependent manner. However, unsaturated C18 and C20 fatty acids (oleate, linolate, linolenate and arachidonate) at 1 mmol/l significantly increased the basal release, but none of them suppressed CRH (1 nmol/l)-induced ACTH release. In the presence of caprylate (1 mmol/l), CRH (1 nmol/l)-stimulated increase in cellular calcium ion concentration was diminished. From these results we conclude that saturated fatty acids have a suppressing effect on CRH-induced ACTH increase in primary cultured rat anterior pituitary cells. Citado por R. Peat.
[A28] BRUDER E D BALL D L, 2003. Am J Physiol Regul Integr Comp Physiol. 2003 Jun;284(6):R1631-5. Epub 2003 Apr 10. An oxidized metabolite of linoleic acid stimulates corticosterone production by rat adrenal cells. Bruder ED1, Ball DL, Goodfriend TL, Raff H. Oxidized derivatives of linoleic acid have the potential to alter steroidogenesis. One such derivative is 12,13-epoxy-9- keto-10-(trans)-octadecenoic acid (EKODE). To evaluate the effect of EKODE on corticosterone production, dispersed rat zona fasciculata/reticularis (subcapsular) cells were incubated for 2 h with EKODE alone or together with rat ACTH (0, 0.2, or 2.0 ng/ml). In the absence of ACTH, EKODE (26 microM) increased corticosterone production from 5.3 +/- 2.3 to 14.7 +/- 5.0 ng. 10(6) cells. h(-1). The stimulatory effect of ACTH was increased threefold in the presence of EKODE (26.0 microM). Cholesterol transport/P-450scc activity was assessed by measuring basal and cAMP-stimulated pregnenolone production in the presence of cyanoketone (1.1 microM). EKODE (13.1 and 26.0 microM) significantly increased basal and cAMP-stimulated (0.1 mM) pregnenolone production. In contrast, EKODE decreased the effect of 1.0 mM cAMP. EKODE had no effect on early or late-pathway activity in isolated mitochondria. We conclude that EKODE stimulates corticosterone biosynthesis and amplifies the effect of ACTH. Increased levels of fatty acid metabolites may be involved in the increased glucocorticoid production observed in obese humans.
[A29] HILLERED, L, CHAN P H, 1989. J Neurosci Res . 1989 Oct;24(2):247-50. doi: 10.1002/jnr.490240216. Brain mitochondrial swelling induced by arachidonic acid and other long chain free fatty acids. L Hillered 1, P H Chan Polyunsaturated fatty acids (PUFAs), arachidonic acid in particular, are well known, potent inducers of edema in the brain, while monounsaturated and saturated long chain fatty acids do not possess this quality. This investigation has compared the ability of some free fatty acids (FFAs), known to be released during cerebral ischemia, to induce brain mitochondrial swelling in vitro. The PUFAs tested, especially arachidonic acid (20:4), were more potent in causing swelling than saturated or monounsaturated ones, as measured by the decrease in light absorbance of the mitochondrial suspension. This finding is in line with the unique potency of 20:4 to induce brain edema. Incubation of brain mitochondria with 20:4 for 20 min caused a dose-dependent swelling. [...] https://pubmed.ncbi.nlm.nih.gov/2531232/
[A30] HILLERED L, CHAN P H, 1998. Role of arachidonic acid and other free fatty acids in mitochondrial dysfunction in brain ischemia. Hillered L, Chan PH.J Neurosci Res. 1988 Aug;20(4):451-6. doi: 10.1002/jnr.490200407.PMID: 3141627
[A31] CHAN P H, FISHMAN R A, 1985. Cellular and molecular effects of polyunsaturated fatty acids in brain ischemia and injury. Chan PH, Fishman RA, Longar S, Chen S, Yu A.Prog Brain Res. 1985;63:227-35. doi: 10.1016/S0079-6123(08)61986-X.PMID: 3939039 Review. No abstract available.
[A32] PEAT, Ray, 2006. Suitable fats, unsuitable fats: issues in nutrition. In: raypeat.com/articles/
[A33] PEAT, Ray, 2013. Fats, functions and malfunctions. http://raypeat.com/articles/articles/fats-functions-malfunctions.shtml
[A34] RUSYN I, 1999. Carcinogenesis. 1999 Nov;20(11):2095-100. Corn oil rapidly activates nuclear factor-kappaB in hepatic Kupffer cells by oxidant-dependent mechanisms. Rusyn I, Bradham CA, Cohn L, Schoonhoven R, Swenberg JA, Brenner DA, Thurman RG.
[A35] ANTHONY M, 1978. Res Clin Stud Headache 1978;6:110-6. Role of individual free fatty acids in migraine. Anthony M “Total plasma free fatty acids, platelet serotonin content and plasma stearic, palmitic, oleic and linoleic acids were estimated in 10 migraine patients before, during and after a migraine attack. Total and individual plasma free fatty acid levels rose and platelet serotonin content fell in most patients. The highest rise was observed in linoleic acid, which is known to be a potent liberator of platelet serotonin in vitro and is the only precursor of all prostaglandins in the body. It is suggested that the rise in plasma levels of linoleic acid in migraine could be responsible for the platelet serotonin release observed during the attack.” Citado por R. Peat.
[A36] GARCIA M C, KIM H Y, 1997. Brain Res 1997 Sep 12;768(1-2):43-8. Mobilization of arachidonate and docosahexaenoate by stimulation of the 5-HT2A receptor in rat C6 glioma cells. Garcia MC, Kim HY Laboratory of Membrane Biochemistry and Biophysics, National Institute on Alcohol Abuse and Alcoholism, National Institutes of Health, Rockville, MD 20852, USA. “In this study, we demonstrate that astroglial 5-HT2A receptors are linked to the mobilization of polyunsaturated fatty acids (PUFA). [...] “These results indicate that the 5-HT2A receptor is coupled to the mobilization of PUFA.” Citado por R. Peat.
[A37] KUJALOVA V, 1975. Bratisl Lek Listy 1975 Jul;64(1):58-63. [The effect of serotonin on the release of free fatty acids from human and rat adipose tissue (author's transl)]. [Article in Czech] Rath R, Kujalova V.
[A38] YOSHIDA, K ASAOKA Y, 1992. Proc Natl Acad Sci U S A 1992 Jul 15;89(14):6443-6. Platelet activation by simultaneous actions of diacylglycerol and unsaturated fatty acids. Yoshida K, Asaoka Y, Nishizuka Y “Several cis-unsaturated fatty acids such as oleic, linoleic, linolenic, eicosapentaenoic, and docosahexaenoic acids added directly to intact human platelets greatly enhance protein kinase C activation as judged by the phosphorylation of its specific endogenous substrate, a 47-kDa protein.” “In the presence of ionomycin and either 1,2-dioctanoylglycerol or phorbol 12-myristate 13-acetate, the release of serotonin from the platelets is also remarkably increased by cis-unsaturated fatty acids. The effect of these fatty acids is observed at concentrations less than 50 microM. Saturated fatty acids and trans-unsaturated fatty acids are inactive.” “. . . cis-unsaturated fatty acids increase an apparent sensitivity of the platelet response to Ca2+. The results suggest that cis-unsaturated fatty acids, which are presumably produced from phosphatidylcholine by signal-dependent activation of phospholipase A2, may take part directly in cell signaling through the protein kinase C pathway.” Citado por R. Peat.
[A39] TAPPIA W J, 1995. Influence of unsaturated fatty acids on the production of tumour necrosis factor and interleukin-6 by rat peritoneal macrophages, Mol. Cell Biochem. 143(2), 89-98, 1995.
[A40] VANPAPENDORP D.H. et al. 1995. , Biochemical profile of osteoporotic patients on essential fatty supplementation, Nutr. Res. 15(3), 325-334, 1995. (Fish oil increased urinary calcium fish oil/evening primrose oil increased osteocalcin and procollagen.)
[A41] FELTON C V, et al., 1994. Dietary polyunsaturated fatty acids and composition of human aortic plaques, Lancet 344(8931), 1195-1196, 1994.
[A42] DARMIANI H, et al., 1994. Interferon-gamma and polyunsaturated fatty acids increase the binding of lipopolysaccharide to macrophages, Int. J. Exp. Pathol. 75(5), 363-368, 1994.
[A43] RAFAEL, J, et al., 1984. The effect of essential fatty acid deficiency on basal respiration and function of liver mitochondria in rats, J. Nutr. 114, 255-262, 1984.
[A44] CHAN P.H. FISHMAN, R A, 1978. Brain edema: Induction in cortical slices by polyunsaturated fatty acids, Science 201, 358-369, 1978. “Ibis cellular edema was specific, since neither saturated fatty acids nor a fatty acid containing a single double bond had such effect.” Citado por R. Peat.
[A45] LARSSON B, et al., 1995. Effects of dietary alpha- and gamma-linolenic acids on liver fatty acids, lipid metabolism, and survival in sepsis, Shock 4(l), 11-20, 1995. “Dietary GLA reduced survival from sepsis.”
[A46] CHEMLA D, et al., 1995. Influence of dietary polyunsaturated fatty acids on contractility, inotropy and compliance of isolated rat myocardium, J mol Cell Cardiol 27(8), 1745-1755, 1995. “There was a trend towards a lower peak lengthening velocity at preload in the LC (n-3) group … together with an unchanged peak rate of isometric force decline. This resulted in a significant impairment of the two mechanical indexes testing the load dependence of myocardial relaxation.” See B. Pieske, Circul. 92(5),1169-78. Citado por R. Peat.
[A47] ENDRESEN M.J. et al., 1994. Effects of free fatty acids found increased in women who develop pre-eclampsia on the ability of endothelial cells to produce prostacyclin, oGMP and inhibit platelet aggregation, Scan. J. Clin. Lab. Invest. 54(7), 549-557, 1994. “…levels of circulating fire fatty acids are increased in women who later develop pre-eclampsia long before the clinical onset of the disease.” “…linoleic acid reduced the thrombin-stimulated prostacyclin release by 30-60% oleic acid by 10-30%, wheras palmitic acid had no effect.” “Linoleic acid reduced the endothelial cells’ ability to inhibit platelet aggregation by 10-45%….” Citado por R. Peat.
[A48] D´AQUINO M. et al., 1991. Effect offish oil and coconut oil on antioxidant defence system and lipid peroxidation in rat liver, Free Radical Res. Commun. (Switzerland) 12-13 (1), 147-152, 1991. The rate of lipid peroxidation in isolated microsomes was three-fold higher in rats fed fish oil as compared to rats with coconut oil diet.” “These results suggest that fish oil feeding at an amount compatible with human diet, although decreasing plasma lipids, actually challenges the antioxidant defence system, thus increasing the susceptibility of tissues to free radical oxidative damage.” Citado por R. Peat
[A49] WANG Y.P. 1995. Aspirin inhibits both lipid peroxides and thromboxane in preeclamptic placentas, Free Radical Biol. Med. 18(3), 585-591, 1995.
[A50] BANGUR C.S. 1995. Thyroid hormone treatment alters phospholipid composition and membrane fluidity of rat brain mitochondria, Biochem. J. 305(1),29-32, 1995. (Increases fluidity.)
[A51] SOHAL et al., 1995. Mitochondrial superoxide and hydrogen peroxide generation, protein oxidative damage,and longevity in different species of flies, Free Rad. Biol. & Med. 19(4),499-504, 1995. Cytochrome C oxidase protects against free radical damage. This enzyme depends on thyroid and light. Citado por R. Peat
[A52] ODA E HATADA K, 2005. Int Heart J. 2005 Nov;46(6):975-85. Relationships between serum unsaturated fatty acids and coronary risk factors: negative relations between nervonic acid and obesity-related risk factors. Oda E, Hatada K, Kimura J, Aizawa Y, Thanikachalam PV, Watanabe K. "The objective of the present study was to analyze the relationships between serum USFA and CRF [coronary risk factors]." "Oleic acid (OA), linoleic acid (LA), and eicosapentaenoic acid (EPA) were positively related to coronary risk factors (total CRFS = 2, 3, and 4, respectively), while nervonic acid (NA) exerted negative effects on these risk factors (total CRFS = -6 ). It is concluded NA may have preventive effects on obesity-related metabolic disorders."
[A53] OMURA M, 2001. FEBS Lett. 2001 Jan 5;487(3):361-6. Eicosapentaenoic acid (EPA) induces Ca(2+)-independent activation and translocation of endothelial nitric oxide synthase and endothelium-dependent vasorelaxation. Omura M, Kobayashi S, Mizukami Y, Mogami K, Todoroki-Ikeda N, Miyake T, Matsuzaki M. "EPA stimulated NO production even in endothelial cells in situ loaded with a cytosolic Ca(2+) chelator . . . which abolished the [Ca(2+)]i elevations induced by ATP and EPA."
[A54] TSOUTSIKOS P MINERS J O, 2004. Biochem Pharmacol. 2004 Jan 1;67(1):191-9. Evidence that unsaturated fatty acids are potent inhibitors of renal UDP-glucuronosyltransferases (UGT): kinetic studies using human kidney cortical microsomes and recombinant UGT1A9 and UGT2B7. Tsoutsikos P, Miners JO, Stapleton A, Thomas A, Sallustio BC, Knights KM.
[A55] AYRE K J HULBERR A J, 1997. Lipids. 1997 Dec;32(12):1265-70. Dietary fatty acid profile affects endurance in rats. Ayre KJ, Hulbert AJ. "The diets comprised an essential fatty acid-deficient diet (containing mainly saturated fatty acids); a diet high in n-6 fatty acids, High n-6; and a diet enriched with n-3 fatty acids, High n-3. Submaximal endurance in rats fed the High n-3 diet was 44% less than in rats fed the High n-6 diet (P < 0.02). All rats were then fed a standard commercial laboratory diet for a 6-wk recovery period, and their performances were reevaluated. Although endurance in all groups was lower then at 9 wk, it was again significantly 50% lower in the High n-3 group than the High n-6 group (P < 0.005). Although n-3 fats are considered beneficial for cardiovascular health, they appear to reduce endurance times, and their side effects need to be further investigated." Citado por R. Peat.
[A56] BARTFAI E, ORSIERE T, 2000. Ann Biol Clin (Paris) 2000 Sep-Oct;58(5):595-600. [Studies on the genotoxic effects of crude liver oils from 3 species of Mediterranean sharks by means of in vitro micronucleus test using human lymphocytes] Bartfai E, Orsiere T, Duffaud F, Villani P, Pompili J, Botta A. "The results of this experimental study show that the crude liver oils of three species of sharks are genotoxic and confirm a high carcinogenic risk." Citado por R. Peat.
[A57] DHEIN S MICHAELIS B, 2005. Naunyn Schmiedebergs Arch Pharmacol. 2005 Mar;371(3):202-11. Epub 2005 Apr 15. Antiarrhythmic and electrophysiological effects of long-chain omega-3 polyunsaturated fatty acids. Dhein S, Michaelis B, Mohr FW. “Atrioventricular conduction time was slowed only by DHA and EPA.” “Regarding antiarrhythmic activity we found that the threshold for elicitation of a ventricular extrasystole was concentration-dependently enhanced by DHA and EPA, but not by ALA. DHA dose-dependently reduced longitudinal propagation velocity V(L) and to a lower extent transverse velocity V(T).”
[A58] DIAZ O BERQUAND A, 2002. J Biol Chem. 2002 Oct 18;277(42):39368-78. The mechanism of docosahexaenoic acid-induced phospholipase D activation inhuman lymphocytes involves exclusion of the enzyme from lipid rafts. Diaz O, Berquand A, Dubois M, Di Agostino S, Sette C, Bourgoin S, Lagarde M, Nemoz G, Prigent AF. “Docosahexaenoic acid (DHA), an n-3 polyunsaturated fatty acid that inhibits T lymphocyte activation, has been shown to stimulate phospholipase D (PLD) activity in stimulated human peripheral blood mononuclear cells (PBMC).” “This PLD activation might be responsible for the immunosuppressive effect of DHA because it is known to transmit antiproliferative signals in lymphoid cells.” Citado por R. Peat
[A59] GAIVA M H, COUTO R C, 2003. Nutrition. 2003 Feb;19(2):144-9. Diets rich in polyunsaturated fatty acids: effect on hepatic metabolism in rats. Gaiva MH, Couto RC, Oyama LM, Couto GE, Silveira VL, Ribeiro EB, Nascimento CM. “Male Wistar rats, just weaned, were fed ad libitum for 8 wk with one of the following diets: rat chow (C), rat chow containing 15% (w/w) soybean oil (S), rat chow containing 15% (w/w) fish oil (F), and rat chow containing 15% soy bean and fish oil (SF; 5:1, w/w).” “Body weight gain was higher in F and SF than in C and S rats. Liver weight, lipid content, and lipogenesis rate increased in F and SF rats, although adenosine triphosphate citrate lyase activity decreased. Glycogen concentration decreased in S, F, and SF rats compared with C rats.” Citado por R. Peat.
[A60] GAIVA M H COUTO R C, 2001. Br J Nutr. 2001 Sep;86(3):371-7. Polyunsaturated fatty acid-rich diets: effect on adipose tissue metabolism in rats. Gaiva MH, Couto RC, Oyama LM, Couto GE, Silveira VL, Riberio EB, Nascimento CM. “Wistar rats were fed ad libitum, for 8 weeks with one of the following diets: C, rat chow; S, rat chow containing 15 % (w/w) soyabean oil; F, rat chow containing 15 % (w/w) fish oil; SF, rat chow containing 15 % (w/w) soyabean and fish oil (5:1, w/w).” "Energy intake was reduced while carcass lipid content was increased in the three fat-fed groups." "These results indicate that enrichment of the diet with polyunsaturated fatty acids causes changes in adipose tissue metabolism that favour fat deposition. Different metabolic pathways were preferentially affected by each type of fatty acid used."
[A61] HARMAN D, 1989. Adv Exp Med Biol 266:3-15, 1989, Lipofuscin and ceroid formation: the cellular recycling system. Harman, D.
[A62] HOUSSAY B A, 1947. B. A. Houssay and C. Martinez, Experimental diabetes and diet, Science 105, 548-549, 1947.
[A63] HULBERT A J, 2005. J Theor Biol. 2005 May 21;234(2):277-88. On the importance of fatty acid composition of membranes for aging. Hulbert AJ. Mech Ageing Dev. 2006 Apr 16; Extended longevity of wild-derived mice is associated with peroxidation-resistant membranes. Hulbert AJ, Faulks SC, Harper JM, Miller RA, Buffenstein R. "Muscle and liver phospholipids from these long-living mice lines have a reduced amount of the highly polyunsaturated omega-3 docosahexaenoic acid compared to the DC mice, and consequently their membranes are less likely to peroxidative damage. The relationship between maximum longevity and membrane peroxidation index is similar for these mice lines as previously observed for mammals in general. It is suggested that peroxidation-resistant membranes may be an important component of extended longevity."
[A64] KAJIHARA H TOTOVIC V, 1975. Virchows Arch B Cell Pathol. 1975 Nov 21;19(3):239-54. [Ultrastructure and morphogenesis of ceroid pigment. II. Late changes of lysosomes in Kupffer cells of rat liver after phagocytosis of unsaturated lipids] Kajihara H, Totovic V, Gedigk P. "These lipids, which have been changed in their molecular structure, cannot be hydrolized by lysosomal enzymes. They remain as an indigestible material, as a waste product in lysosomal residual bodies. Both lipofuscin and ceroid are lysosomal structures containing oxidized and polymerized lipids."
[A65] Free Radic Biol Med 1999 Feb;26(3-4):260-5. Modulation of cardiac mitochondrial membrane fluidity by age and calorie intake. Lee J, Yu BP, Herlihy JT. "The fatty acid composition of the mitochondrial membranes of the two ad lib fed groups differed: the long-chain polyunsaturated 22:4 fatty acid was higher in the older group, although linoleic acid (18:2) was lower. DR eliminated the differences." "Considered together, these results suggest that DR maintains the integrity of the cardiac mitochondrial membrane fluidity by minimizing membrane damage through modulation of membrane fatty acid profile."
[A66] MARTINEZ M BALLABRIGA A, 1987. Lipids 22(3), 133-6, 1987. Effects of parenteral nutrition with high doses of linoleate on the developing human liver and brain, Martinez, M., and A. Ballabriga.
[A67] MASCOLO N, IZZO A A, 1995. J Pharmacol Exp Ther. 1995 Jan;272(1):469-75. Acetic acid-induced colitis in normal and essential fatty acid deficient rats. Mascolo N, Izzo AA, Autore G, Maiello FM, Di Carlo G, Capasso F.
[A68] FEBS Lett. 1998 Oct 16;437(1-2):24-8. Generation of protein carbonyls by glycoxidation and lipoxidation reactions with autoxidation products of ascorbic acid and polyunsaturated fatty acids. Miyata T, Inagi R, Asahi K, Yamada Y, Horie K, Sakai H, Uchida K, Kurokawa K. Citado por R. Peat.
[A69] REITHMANN C, 1996. Naunyn Schmiedebergs Arch Pharmacol. 1996 Jul;354(2):109-19. Exposure to the n-3 polyunsaturated fatty acid docosahexaenoic acid impairs alpha 1-adrenoceptor-mediated contractile responses and inositol phosphate formation in rat cardiomyocytes. Reithmann C, Scheininger C, Bulgan T, Werdan K. “The results presented show that chronic n-3 polyunsaturated fatty acid pretreatment of rat cardiomyocytes leads to a marked impairment of alpha 1-adrenoceptor-induced positive inotropic effects and induction of arrhythmias concomitant with a n-3 fatty acid-induced decrease in IP3 formation.” Citado por R. Peat
[A70] SAKAMORO, N NISHIIKE, T, 2000. Nutrition. 2000 Jan;16(1):11-4. Effects of eicosapentaenoic acid intake on plasma fibrinolytic and coagulation activity by using physical load in the young. Sakamoto N, Nishiike T, Iguchi H, Sakamoto K. “Thus, as determined by the load, a small amount of daily EPA intake clearly decreased fibrinolytic activity and increased coagulation activity.” Citado por R. Peat
[A71] SASAKI T, 1998. Nutr Cancer 1998;30(2):137-43. Effects of dietary n-3-to-n-6 polyunsaturated fatty acid ratio on mammary carcinogenesis in rats. Sasaki T, Kobayashi Y, Shimizu J, Wada M, In'nami S, Kanke Y, Takita T. "Dietary fat was fed to the rats as 10% of the total feed weight, starting two weeks before the initiation. An increase in the n-3/n-6 ratio did not suppress the incidence or reduce the latency of mammary tumor development. The number and weight of mammary tumors per tumor-bearing rat tended to be large in the group with an n-3/n-6 ratio of 7.84 compared with those in the other groups. As the n-3/n-6 ratios were elevated, the total number and weight of tumors increased gradually." "These results suggested that the increase in the n-3/n-6 ratio of dietary fat with the fixed PUFA-to-saturated fatty acid ratio cannot suppress the mammary carcinogenesis but can promote development of tumors, despite reduced PGE2 concentration in the tumor." Citado por R. Peat.
[A72] SONG J H FUJIMOTO K, 2000. J Nutr 2000 Dec;130(12):3028-33. Polyunsaturated (n-3) fatty acids susceptible to peroxidation are increased in plasma and tissue lipids of rats fed docosahexaenoic acid-containing oils. Song JH, Fujimoto K, Miyazawa T.. "Thus, high incorporation of (n-3) fatty acids (mainly DHA) into plasma and tissue lipids due to DHA-containing oil ingestion may undesirably affect tissues by enhancing susceptibility of membranes to lipid peroxidation and by disrupting the antioxidant system."
[A73] STEERENBERG P A, 2002. Diabetes Nutr Metab. 2002 Aug;15(4):205-14. Long-term effect of fish oil diet on basal and stimulated plasma glucose and insulin levels in ob/ob mice. Steerenberg PA, Beekhof PK, Feskens EJ, Lips CJ, Hoppener JW, Beems RB. “We have investigated, in comparison to low and high fat diets, the effect of a fish oil diet on basal and stimulated plasma glucose and insulin levels in male and female ob/ob mice.” “Intercurrent deaths were found especially in the fish oil diet group. Compared to the other diet groups, plasma insulin levels of the fish oil diet group were significantly increased 3 months after the start of the diet and remained higher for another 3 months.” “At 12 months, microscopy revealed an increased severity of hepatic brown pigment accumulation and extramedullary haematopoiesis in the spleen of mice fed with fish oil.” “Fish oil diet also increased intercurrent mortality. However, a consistent course of death could not be established using morphological parameters.” Citado por R. Peat
[A74] WANG Y, 2002. J Biol Chem. 2002 Feb 15;277(7):5692-7. Unsaturated fatty acids inhibit cholesterol efflux from macrophages by increasing degradation of ATP-binding cassette transporter A1. Wang Y, Oram JF. “These findings raise the possibility that an increased supply of unsaturated fatty acids in the artery wall promotes atherogenesis by impairing the ABCA1 cholesterol secretory pathway in macrophages.” Citado por R. Peat
[A75] WANG Y, 2005. J Biol Chem. 2005 Oct 28;280(43):35896-903. Epub 2005 Aug 23. Unsaturated fatty acids phosphorylate and destabilize ABCA1 through a phospholipase D2 pathway. Wang Y, Oram JF. "ATP-binding cassette transporter ABCA1 mediates the transport of cholesterol and phospholipids from cells to HDL apolipoproteins and thus modulates HDL levels and atherogenesis. Unsaturated fatty acids, which are elevated in diabetes, impair the ABCA1 pathway in cultured cells by destabilizing ABCA1 protein." "Unsaturated but not saturated fatty acids stimulated phospholipase D (PLD) activity, the PLD inhibitor 1-butanol prevented the unsaturated fatty acid-induced reduction in ABCA1 levels, and the PLD2 activator mastoparan markedly reduced ABCA1 protein levels, implicating a role for PLD2 in the ABCA1 destabilizing effects of fatty acids." "These data provide evidence that intracellular unsaturated acyl-CoA derivatives destabilize ABCA1 by activating a PLD2 signaling
[A76] ALVAREZ-GUERRA, M, HANNAERT P, 2003. Eur J Pharmacol. 2003 Apr 11;466(1-2):199-205. Vascular permeabilization by intravenous arachidonate in the rat peritoneal cavity: antagonism by antioxidants. Alvarez-Guerra M, Hannaert P, Hider H, Chiavaroli C, Garay RP.
[A77] ANDZIAK B, 2006. Aging Cell. 2006 Dec;5(6):525-32. Disparate patterns of age-related changes in lipid peroxidation in long-lived naked mole-rats and shorter-lived mice. Andziak B, Buffenstein R.
[A78] CICALA G, 1994, J Lipid Mediat Cell Signal. 1994 Mar;9(2):145-53. Essential fatty acid-deficient diet modifies PAF levels in stomach and duodenum of endotoxin-treated rats. Autore G, Cicala C, Cirino G, Maiello FM, Mascolo N, Capasso F.
[A79] BENASSAYAG C, 1999. Prostaglandins Leukot Essent Fatty Acids. 1999 May-Jun;60(5-6):393-9. Does high polyunsaturated free fatty acid level at the feto-maternal interface alter steroid hormone message during pregnancy? Benassayag C, Rigourd V, Mignot TM, Hassid J, Leroy MJ, Robert B, Civel C, Grange' G, Dallot E, Tanguy J, Nunez EA, Ferre' F.
[A80] CHILDS, C E, 2008. Proc Nutr Soc. 2008 Feb;67(1):19-27. Gender differences in the n-3 fatty acid content of tissues. Childs CE, Romeu-Nadal M, Burdge GC, Calder PC.
[A81] DOMBROWSKY H, 2011. J Nutr. 2011 Sep;141(9):1635-42. Ingestion of (n-3) fatty acids augments basal and platelet activating factor-induced permeability to dextran in the rat mesenteric vascular bed. Dombrowsky H, Lautenschläger I, Zehethofer N, Lindner B, Schultz H, Uhlig S, Frerichs I, Weiler N.
[A82] DURAND G, 1980. Ann Nutr Aliment. 1980;34(2):317-32. [Polyunsaturated fatty acids and aging. Lipofuscins : structure, origin and development] [Article in French] Durand G, Desnoyers F.
[A83] J Physiol. 1998 Mar 1;507 ( Pt 2):541-7. Arachidonic acid increases cerebral microvascular permeability by free radicals in single pial microvessels of the anaesthetized rat. Easton AS, Fraser PA.
[A84] GILTAY E J, 2004. Am J Clin Nutr. 2004 Nov;80(5):1167-74. Docosahexaenoic acid concentrations are higher in women than in men because of estrogenic effects. Giltay EJ, Gooren LJ, Toorians AW, Katan MB, Zock PL.
[A85] HOCH F L, 1992. Biochim Biophys Acta. 1992 Mar 26;1113(1):71-133. Cardiolipins and biomembrane function. Hoch FL.
[A86] IRVINE R F, 2002. Sci STKE. 2002 Sep 17;2002(150):re13. Nuclear lipid signaling. Irvine RF.
[A87] LEE H J, 2006. Lipids Health Dis. 2006 Jan 23;5:2. Selective remodeling of cardiolipin fatty acids in the aged rat heart. Lee HJ, Mayette J, Rapoport SI, Bazinet RP.
[A88] MITCHELL T W, 2007. Exp Gerontol. 2007 Nov;42(11):1053-62. Membrane phospholipid composition may contribute to exceptional longevity of the naked mole-rat (Heterocephalus glaber): a comparative study using shotgun lipidomics. Mitchell TW, Buffenstein R, Hulbert AJ.
[A89] NANJI AUTOR, 2001. J Pharmacol Exp Ther. 2001 Nov;299(2):638-44. Dietary saturated fatty acids reverse inflammatory and fibrotic changes in rat liver despite continued ethanol administration. Nanji AA, Jokelainen K, Tipoe GL, Rahemtulla A, Dannenberg AJ.
[A90] NOUROOZ-ZADEH, 1999. Ophthalmic Res. 1999;31(4):273-9. Age-related accumulation of free polyunsaturated fatty acids in human retina. Nourooz-Zadeh J, Pereira P.
[A91] OARADA M, FURUKAWA H, 2000. Biochim Biophys Acta. 2000 Aug 24;1487(1):1-14. Fish oil diet affects on oxidative senescence of red blood cells linked to degeneration of spleen cells in mice. Oarada M, Furukawa H, Majima T, Miyazawa T.
[A92] PARADIES G, 1997. FEBS Lett. 1997 Apr 7;406(1-2):136-8. Age-dependent decline in the cytochrome c oxidase activity in rat heart mitochondria: role of cardiolipin. Paradies G, Ruggiero FM, Petrosillo G, Quagliariello E.
[A93] PERSAD R A, 1990. Br J Urol. 1990 Mar;65(3):268-70. Erythrocyte stearic to oleic acid ratio in prostatic carcinoma. Persad RA, Gillatt DA, Heinemann D, Habib NA, Smith PJ.
[A94] PRASAD A, 2010. J Cell Physiol. 2010 Nov;225(3):829-36. Role of calcium and ROS in cell death induced by polyunsaturated fatty acids in murine thymocytes. Prasad A, Bloom MS, Carpenter DO.
[A95] RAFAEL J, 1984. J Nutr. 1984 Feb;114(2):255-62. The effect of essential fatty acid deficiency on basal respiration and function of liver mitochondria in rats.Rafael J, Patzelt J, Schäfer H, Elmadfa I.
[A96] SINGH R K, 1995. Invasion Metastasis. 1995;15(3-4):144-55. Stearate inhibits human tumor cell invasion. Singh RK, Hardy RW, Wang MH, Williford J, Gladson CL, McDonald JM, Siegal GP.
[A97] SOLFRIZZI V, 2005. Exp Gerontol. 2005 Apr;40(4):335-43. Unsaturated fatty acids intake and all-causes mortality: a 8.5-year follow-up of the Italian Longitudinal Study on Aging. Solfrizzi V, D'Introno A, Colacicco AM, Capurso C, Palasciano R, Capurso S,Torres F, Capurso A, Panza F.
[A98] SPEAKE B K, 1997. Biochim Biophys Acta. 1997 Apr 21;1345(3):317-26. The preferential mobilisation of C20 and C22 polyunsaturated fatty acids from the adipose tissue of the chick embryo: potential implications regarding the provision of essential fatty acids for neural development. Speake BK, Cerolini S, Maldjian A, Noble RC.
[A99] STRIPP C, 2003. J Nutr. 2003 Nov;133(11):3664-9. Fish intake is positively associated with breast cancer incidence rate. Stripp C, Overvad K, Christensen J, Thomsen BL, Olsen A, Moller S, Tjonneland A.
[A100] THOMSON A B, 1989. Am J Physiol. 1989 Jan;256(1 Pt 1):G178-87. Saturated fatty acid diet prevents radiation-associated decline in intestinal uptake. Thomson AB, Keelan M, Lam T, Cheeseman CI, Walker K, Clandinin MT.
[A101] TSOUTSIKOS P, 2004. Biochem Pharmacol. 2004 Jan 1;67(1):191-9. Evidence that unsaturated fatty acids are potent inhibitors of renal UDP-glucuronosyltransferases (UGT): kinetic studies using human kidney cortical microsomes and recombinant UGT1A9 and UGT2B7.Tsoutsikos P, Miners JO, Stapleton A, Thomas A, Sallustio BC, Knights KM.
[A102] YANG D Y, PAN H C, 2007. Neurotoxicology. 2007 Nov;28(6):1220-9. Detrimental effects of post-treatment with fatty acids on brain injury in ischemic rats. Yang DY, Pan HC, Yen YJ, Wang CC, Chuang YH, Chen SY, Lin SY, Liao SL, Raung SL, Wu CW, Chou MC, Chiang AN, Chen CJ.
***
ATENÇÃO!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!! ESTE BLOG NÃO ESTÁ MAIS PUBLICANDO ARTIGOS NOVOS.
POR MOTIVO DE FORÇA MAIOR.
SOLICITAMOS, POR GENTILEZA, AOS INSCRITOS NESTE BLOG QUE MIGREM E SE INSCREVAM NO NOVO BLOG QUE É, NA VERDADE UMA CONTINUAÇÃO DESTE.
O NOVO BLOG, TAMBÉM NO SUBSTACK,
E QUE SUBSTITUI ESTE,
SE CHAMA OUTRAMEDICINA2024.
Pode ser acessado aqui: https://outramedicina2024.substack.com/